
Methylene Blue vs CO₂
Artificial Shortcut or Foundational Restoration?
Something interesting is happening in longevity and biohacking circles. A compound that was once confined to hospital formularies and chemistry labs has found its way into the morning routines of people searching for sharper minds and more resilient bodies. That compound is methylene blue and the reports surrounding it are difficult to ignore.
People who take it describe increased mental clarity, more sustained physical energy, better tolerance for stress, and a quality of focus that feels qualitatively different from caffeine or other common stimulants. These are not fringe anecdotes. They are consistent enough to demand a serious physiological explanation.
But to understand what methylene blue is actually doing, you first have to understand the deeper system it is interacting with a system most people know almost nothing about. And once you understand that system, a more important question emerges: not whether methylene blue works, but why so many people need it in the first place.
The Body Runs on Electron Flow
Most people understand the body in terms of calories energy consumed, energy burned. But this framing misses the actual biochemical mechanism entirely. The body does not run on calories. It runs on electron flow.
When food is broken down through metabolism, what is really happening is that high-energy electrons are being extracted from fuel molecules and loaded onto carrier compounds inside the cell. These carriers primarily NADH and FADH₂ then transport those electrons into the mitochondria, where they enter a precisely organized sequence of protein complexes called the electron transport chain.
As electrons move through this chain from Complex I through CoQ10, Complex III, cytochrome c, and finally Complex IV they drive the pumping of protons across the inner mitochondrial membrane. That proton gradient creates an electrochemical potential, a kind of molecular voltage, which ATP synthase then uses to manufacture ATP. ATP is the energy currency that powers every heartbeat, every nerve impulse, every immune response, every thought. The entire economy of life depends on this one process running smoothly.
The electron transport chain is not merely an energy system. It is simultaneously an electrical system, a redox system, an oxygen utilization system, and a proton gradient system.
When electrons stop flowing efficiently, the consequences are not isolated. Because the Krebs cycle requires NAD⁺ and FAD the oxidized forms of those carrier molecules to continue operating, any slowdown in the ETC causes those carriers to accumulate in their reduced state. NAD⁺ declines. The Krebs cycle stalls. ATP production falls. Reactive oxygen species begin leaking from the chain. Oxidative stress rises. The dysfunction spreads upstream like a traffic jam backing up from a blocked highway.
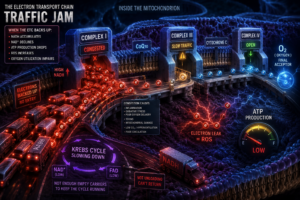
What Slows Electron Flow
There is no single cause. Electron flow can be compromised by poor oxygen delivery, impaired microcirculation, chronic inflammation, mitochondrial membrane damage, environmental toxins, and ischemia. But one factor sits at the center of modern mitochondrial dysfunction more than almost any other, and it receives almost no attention: carbon dioxide.
CO₂ is universally treated as a waste gas. The body makes it, the lungs remove it, and that is the end of the conversation. This framing is not merely incomplete it is physiologically backwards. Carbon dioxide is one of the most important regulatory molecules in the body, and its role in oxygen delivery alone makes it central to mitochondrial function.
The mechanism is called the Bohr Effect. Hemoglobin the molecule that carries oxygen through the bloodstream does not release oxygen at a fixed rate. Its affinity for oxygen is regulated by the local CO₂ concentration. When CO₂ is adequate, hemoglobin releases oxygen efficiently into the tissue that needs it. When CO₂ is low, hemoglobin holds onto oxygen more tightly, and the delivery to tissue worsens.
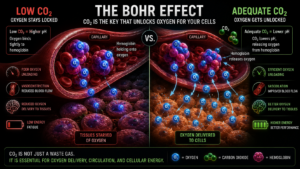
The result is a situation that appears paradoxical but is physiologically real: a person can have normal blood oxygen saturation on a pulse oximeter while their cells are simultaneously starved for the oxygen that is sitting unused in their bloodstream. This is one of the most clinically underappreciated gaps in modern medicine. Standard blood panels measure whether oxygen is present in circulation. They do not measure whether oxygen is actually reaching the mitochondria. A reading of 98% saturation tells you hemoglobin is loaded. It tells you nothing about whether that oxygen is being released at the tissue level where it is needed.
This matters enormously for understanding why people with “normal” labs can feel profoundly unwell. Brain fog, fatigue that does not respond to sleep, exercise intolerance, cold hands and feet, poor recovery these are not mysterious symptoms without a mechanism. They are exactly what you would expect from a system in which oxygen is being transported but not delivered. The mitochondria are waiting at the end of the line, and the delivery never arrives.
Normal blood oxygen saturation does not mean oxygen is being delivered. It means oxygen is being carried. Those are not the same thing.
CO₂ also acts as a primary pH buffer and a vasodilator. When carbon dioxide falls whether through chronic overbreathing, anxiety, poor posture, or mouth breathing blood vessels constrict, particularly in the brain and microcirculation. This is not a minor effect. Studies of cerebral blood flow show meaningful reductions in brain perfusion even at modest drops in CO₂, which is why hyperventilation rapidly produces dizziness, cognitive blurring, and a sense of unreality. The brain is not getting less oxygen in the blood. It is getting less blood altogether.
Respiratory alkalosis develops alongside this vasoconstriction. The proton balance the mitochondria depend on for their electrochemical gradients begins to shift. And because the electron transport chain is exquisitely sensitive to both pH and membrane potential operating within tolerances that are measured in fractions of a pH unit even moderate alkalosis begins destabilizing the chain. Electron flow slows. Reactive oxygen species increase. The system that was supposed to be protected by adequate oxygen delivery becomes less able to use the oxygen that does arrive.
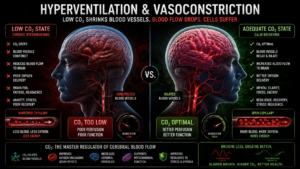
What makes this so insidious is that none of it shows up on standard diagnostic tests. CO₂ tolerance is not routinely measured. Microcirculatory function is not routinely assessed. Mitochondrial efficiency is not something a standard metabolic panel captures. The dysfunction accumulates invisibly, and the person is left with symptoms they cannot explain and results that tell them nothing is wrong.
This is the terrain methylene blue steps into.
How Methylene Blue Acts as a Bypass
Methylene blue is what chemists call a redox mediator. It can be reduced accepting electrons and then oxidized again donating them. This reversibility is what makes it pharmacologically useful inside the mitochondria.
Under normal conditions, electrons move through the ETC in a highly coordinated sequence. When congestion develops when electrons back up behind a bottleneck in the chain methylene blue can step in as an auxiliary shuttle. It accepts electrons upstream of the blockage, converts into its reduced form called leucomethylene blue, then travels further downstream toward cytochrome c and Complex IV, where it donates those electrons back into the chain. At that point it oxidizes again, back into methylene blue, ready to repeat the cycle.
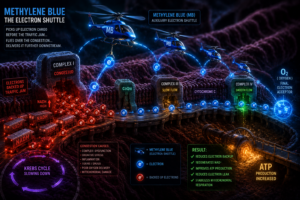
The downstream effects of this bypass are real and measurable. NAD⁺ is regenerated, allowing the Krebs cycle to continue. Electron leak is reduced, which means fewer reactive oxygen species escaping into the cell. ATP production improves. Mitochondrial respiration stabilizes. This is why the reported effects of methylene blue are not imaginary. The molecule is doing genuine biochemical work.
But here is the question that should follow every honest account of what methylene blue does: what caused the traffic jam in the first place? The bypass is real. The relief is real. But a bypass is not repair. And the distinction matters more than almost anything else in this conversation.
The Limitation of the Bypass
Consider the difference between two responses to a blocked highway. The first is a helicopter service that picks up cargo before the blockage, flies over it, and delivers it downstream. The second is a crew that actually clears the road. Both get cargo moving. But only one of them addresses the condition that caused the problem. And critically only one of them means the highway still needs a helicopter tomorrow.
Methylene blue is the helicopter. And the helicopter works. In acute situations neurological stress, severe oxidative damage, mitochondrial crisis, recovery from illness the bypass may be exactly what the system needs to buy time. There are legitimate clinical applications here, and dismissing them would be as intellectually dishonest as overstating them.

But the terrain underneath does not change because the helicopter is flying. If the underlying conditions driving mitochondrial dysfunction persist chronically low CO₂, impaired oxygen delivery, microcirculatory compromise, systemic oxidative stress then the moment the bypass stops, the traffic jam returns. The person has not been restored. They have been rerouted.
There is a physiological cost to permanent compensation that rarely gets discussed. When the body is chronically rerouting electron flow rather than resolving the conditions that impair it, that compensation is not free. The mitochondria are working harder to maintain output through a less efficient pathway. The system is under greater oxidative load because the bypass, while helpful, is messier than the primary chain. Inflammation does not resolve because the root drivers of electron congestion poor oxygen delivery, pH instability, chronic sympathetic activation are still present and still generating the signals that trigger inflammatory cascades.
Over time, this creates a pattern that many people recognize but struggle to name: they feel better when taking methylene blue, noticeably worse when they stop, and never quite reach a baseline that feels genuinely stable. That pattern is not evidence that methylene blue is indispensable. It is evidence that the bypass has become load-bearing that the body has organized itself around the compensation rather than resolving what made the compensation necessary.
This is not a reason to avoid methylene blue. It is a reason to use it with a clear understanding of what it is doing and what it cannot do. A bridge has value. But a bridge is not a destination.
Why CO₂ Is More Foundational
What makes carbon dioxide genuinely different from methylene blue is not simply that it is natural versus synthetic. It is that CO₂ is involved in almost every layer of the problem, and its restoration works with the body’s own regulatory architecture rather than around it.
Healthy mitochondria produce carbon dioxide as a direct byproduct of efficient aerobic metabolism. The Krebs cycle releases CO₂ at multiple steps. This means that CO₂ production is itself a marker of mitochondrial health, not just a consequence of it. And when CO₂ is adequate, it maintains the conditions healthy mitochondria require: stable pH, efficient oxygen unloading, dilated vasculature, stable proton gradients, reduced oxidative stress.
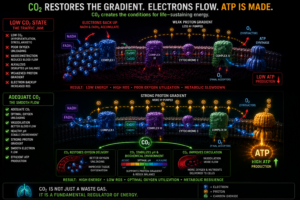
The loop is self-reinforcing in both directions. Healthy mitochondria produce CO₂. Adequate CO₂ maintains the conditions healthy mitochondria need. But when mitochondrial function declines, CO₂ production drops, which worsens oxygen delivery, which further impairs mitochondrial function, which further reduces CO₂.
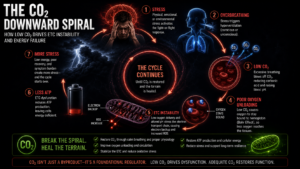
Most modern humans chronically overbreathe. Not dramatically. Subtly, persistently, and with consequences that accumulate slowly enough to become invisible.
The drivers of chronic CO₂ depletion are not exotic. They are the ordinary features of modern life: chronic psychological stress, which accelerates breathing; mouth breathing, which increases ventilation volume; sedentary behavior, which reduces metabolic CO₂ production; and sympathetic nervous system dominance, which keeps the body in a low-grade fight-or-flight state that perpetually up-regulates breathing rate. None of these cause acute catastrophe. Together, over years, they slowly drain the CO₂ reserves the mitochondria depend on.
Restoring CO₂ physiology through breathwork specifically through practices that reduce breathing volume and raise the body’s CO₂ tolerance does not introduce an external compensatory molecule. It restores a regulatory molecule the body already evolved to produce. It rebuilds the road rather than routing cargo around the blockage.
Two Different Questions
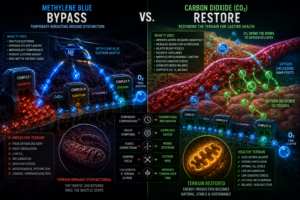
Methylene blue and CO₂ restoration represent fundamentally different therapeutic orientations, and the difference can be stated simply.
Methylene blue asks: how do we bypass the dysfunction? It introduces an external electron shuttle to compensate for a compromised chain. The work is real, the effects are real, and in the right context the approach has genuine value.
CO₂ restoration asks: how do we restore the conditions where the dysfunction becomes less likely to occur? It addresses oxygen delivery, pH stability, proton gradients, vasomotor tone, and mitochondrial stability at their root. The work is slower, less dramatic, and harder to quantify but it aims at the terrain rather than the symptom.
One approach works from the outside in, compensating for a compromised system. The other works from the inside out, restoring the system’s capacity to regulate itself.
This does not make methylene blue illegitimate. There are clinical situations where a bypass is the right call where the system is under enough acute stress that restoring terrain first is not realistic, and a compensatory bridge is needed. In those cases, methylene blue may buy the time and stability that make deeper restoration possible.
But the popularity of methylene blue reveals something important about the state of modern physiology. When a large portion of the population feels dramatically better after introducing an external electron shuttle into their mitochondria, it suggests that electron flow is already compromised on a population scale. That is not a supplement story. That is a civilization story. And the most important question it raises is not which molecule to take, but what conditions have made so many people dependent on the bypass in the first place.
A Note on Caution
Methylene blue is a pharmacologically active compound, not a benign supplement, and it deserves to be treated as one. The casual framing it often receives in biohacking communities as a safe cognitive enhancer you can titrate up freely does not reflect its actual pharmacological profile.
The dose-response relationship is particularly important to understand. At low doses, typically in the range of 0.5 to 2 milligrams per kilogram of body weight, methylene blue acts as an antioxidant and electron shuttle, doing the mitochondrial support work described throughout this article. But the curve does not continue upward linearly. At higher doses, methylene blue flips its mechanism and becomes pro-oxidant generating the very reactive oxygen species it was supposed to reduce. This is not a theoretical concern. It is a well-documented property of the molecule, and it means that more is not better. A person taking a higher dose in search of stronger effects may be pushing the compound past the threshold where it supports mitochondrial function into the range where it begins damaging it.
The interaction risk with serotonergic medications is more serious still. Methylene blue inhibits monoamine oxidase A, the enzyme responsible for breaking down serotonin in the brain. When combined with medications that increase serotonin availability SSRIs, SNRIs, MAOIs, certain migraine medications, and others this inhibition can cause serotonin to accumulate to dangerous levels. Serotonin syndrome ranges in severity from mild symptoms like agitation, rapid heart rate, and muscle twitching to life-threatening presentations including hyperthermia, seizures, and cardiovascular instability. This is not a rare theoretical interaction. It has been documented in clinical settings, including cases involving intravenous methylene blue administered during surgery to patients on antidepressants.
Anyone currently taking psychiatric medications, serotonergic drugs, or medications that affect monoamine metabolism should consider methylene blue off-limits without direct guidance from a physician who understands the interaction profile. The fact that it is available without a prescription and widely discussed as a wellness compound does not reduce this risk. If anything, the casual context increases it by creating a false impression of safety.
Purity also matters. Pharmaceutical-grade methylene blue is a different product from the laboratory-grade chemical reagent that appears in some supply chains. Contaminants in lower-grade preparations can introduce their own toxicity. Anyone using methylene blue should verify they are using a pharmaceutical-grade product from a reputable source.
The Bigger Picture

Carbon dioxide is not simply a waste gas. It is one of the central regulators of human energy production a molecule so deeply woven into the physiology of respiration, circulation, mitochondrial function, and cellular redox balance that its decline touches almost every system in the body simultaneously.
The goal is not to find a better bypass. The goal is to restore the conditions where efficient energy production can occur naturally again where the body does not need a helicopter because the road is clear.
Methylene blue may have a place in that process, particularly during acute phases where the system needs time to stabilize. But the endpoint is not continued dependence on an external electron shuttle. The endpoint is a body that generates its own CO₂, delivers its own oxygen, and runs its own mitochondrial chain without needing compensation.
The practical path toward that endpoint is less exotic than most people expect. It begins with breathing. Not breathing exercises in the dramatic sense not forced breath retention or aggressive techniques but the slow, consistent work of reducing chronic over-breathing. This means learning to breathe through the nose rather than the mouth, which naturally reduces ventilation volume and preserves CO₂. It means recognizing the connection between psychological stress and breathing rate, and developing the capacity to bring both down together. It means building CO₂ tolerance gradually through reduced-volume breathing practices, so that the body stops treating its own carbon dioxide as something to be eliminated as fast as possible and starts treating it as the regulatory molecule it actually is.
None of this is fast. The body’s CO₂ tolerance measured by how long a person can comfortably extend an exhale or tolerate a gentle air hunger reflects years of breathing patterns. Changing it takes consistent practice over weeks and months. But unlike an external supplement, the gains compound. As CO₂ tolerance improves, oxygen delivery improves. As oxygen delivery improves, mitochondrial function stabilizes. As mitochondrial function stabilizes, the body produces more CO₂ naturally. The downward spiral reverses. The self-reinforcing loop begins running in the right direction.
That is restoration. Not rerouting, not compensation, not a bypass that needs to be maintained indefinitely. The return of a system to the conditions it was designed to operate within.
Methylene blue may temporarily reroute the current. But carbon dioxide may help rebuild the electrical grid itself.
learn more at: THECARBONATEDBODY.COM

